ニュース
![]() 2022.12.21
2022.12.21
【研究成果】細胞接着面切り替わりの分子・力学メカニズムを解明
東京大学
名古屋大学
発表者
井川 敬介(東京大学 大学院理学系研究科 生物科学専攻 特任助教(研究当時)/現:名古屋大学 大学院理学研究科 理学専攻 助教)
石原 秀至(東京大学 大学院総合文化研究科 広域科学専攻 准教授)
田守 洋一郎(京都大学 大学院医学研究科 分子生体統御学講座 准教授)
杉村 薫(東京大学 大学院理学系研究科 生物科学専攻 准教授)
発表のポイント
- 細胞の配置換え過程でアドヘレンスジャンクションからミオシンが剥離する現象を足がかりにして、細胞接着面切り替わりの分子・力学メカニズムを明らかにしました。
- アドヘレンスジャンクションの構成因子Jub/Ajubaとトリセルラージャンクションの構成因子M6とが相互に抑制しあうことで、細胞接着面切り替わり時のミオシンの剥離と再接着が制御されることを明らかにしました。
- ミオシンケーブルの張力と細胞接着面への接着との競合を考慮した力学モデルを濡れの理論に基づいて構築し、細胞配置換えにおける細胞接着面の収縮と伸長に伴って、ミオシンの剥離と再接着が自発的に起こるメカニズムを同定しました。
発表概要
生き物の体は、組織の秩序だった変形が繰り返されることで形づくられます。組織の変形を個々の細胞レベルでみると、細胞の数、かたち、配置の変化に分解されます。このうち、細胞接着面の組み替えによる細胞の相対位置の変化は細胞配置換えと呼ばれます(図1a)。細胞配置換えは、細胞接着面の収縮、切り替わり、伸長の三段階で進行します(図1b)。細胞接着面の収縮と伸長はミオシン(注1)が生成する力により駆動されることが分かっていますが、細胞接着面切り替わりのメカニズムには多くの謎が残されていました。 今回、東京大学大学院理学系研究科生物科学専攻の井川敬介特任助教(研究当時)と杉村薫准教授らの研究グループは、細胞接着面の切り替え時にアドへレンスジャンクション(注2)からミオシンが剥離する構造 (rectangle-shaped myo-II cable: rsMC)を発見したことを足がかりに、細胞接着面切り替わりの分子・力学メカニズムを明らかにしました(図2-4)。ショウジョウバエ翅上皮細胞では、アドへレンスジャンクションとアクチンの間のリンカー分子として働くことが知られているJub/Ajubaが、細胞配置換え初期にミオシンが細胞接着面から剥離しないように働きます。加えて、トリセルラージャンクション(注3)の構成因子M6がJub/Ajubaの局在を減弱させることで、rsMCが形成されます。Jub/AjubaとM6は相互抑制関係にあり、両者の相互抑制のバランスとミオシンケーブルの物理特性はともに、細胞接着面の長さに依存しています。このように、幾何と力学、シグナリングが協調することで、細胞配置換えにおける細胞接着面の収縮と伸長に伴って、自発的に、細胞接着面の切り替えが起こることが明らかになりました。 本研究は、長年、ブラックボックスになっていた細胞接着切り替わりのメカニズムの一端を明らかにしました。細胞間接着の動態は生き物の体の維持から老化などのライフイベント、がんなどの病気と深く関わっており、本研究により明らかにされた細胞間接着制御メカニズムは、さまざまな生命現象において働いている可能性があります。
発表内容
[研究の背景]
組織が正しい形に成長するためには、細胞の配置換えが精緻に制御される必要があります。この10年ほどで、細胞配置換えに伴う細胞接着面の収縮と伸長の分子メカニズムの多くが明らかにされました。特に、ミオシンによる力の生成が細胞接着面の収縮と新生細胞接着面の伸長の両方を担うことが分かっています(図1)。一方で、細胞配置換えが達成されるためには、接着する細胞が切り替わる細胞間接着の再編成が必須ですが、そのメカニズムはほとんど明らかにされていませんでした。
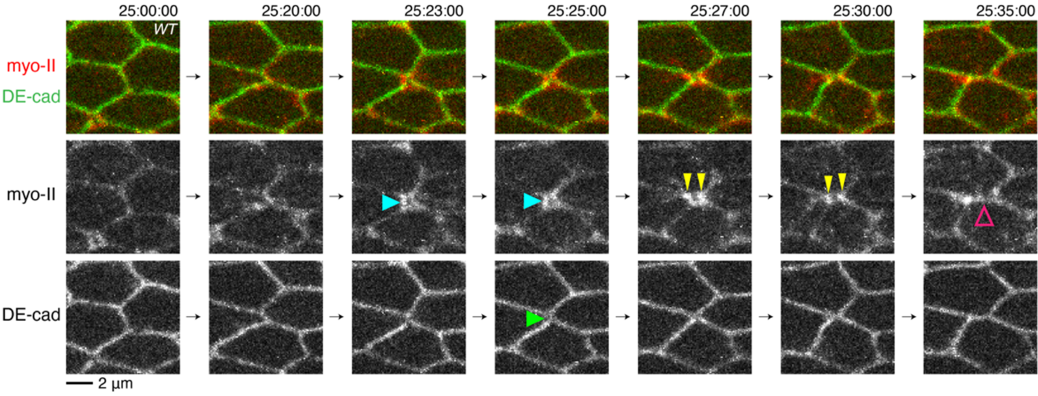
本研究グループは以前、ショウジョウバエ翅上皮細胞の接着面が組み換わる直前に、ミオシンがアドへレンスジャンクションから一過的に剥離し、rsMC (rectangle-shaped myo-II cable) と呼ばれる構造を形成することを見出していました(Ikawa and Sugimura, 2018, Nat. Commun.; 図2a-c)。加えて、rsMCの構造が乱れると、細胞配置換えが不安定化することも分かっていました。これらの観察事実は、rsMCが適切に形成されることが細胞接着面の切り替わりに必須の役割を果たしていると示唆します。しかし、rsMCの構造と動態がどのように制御されているのかは不明でした。
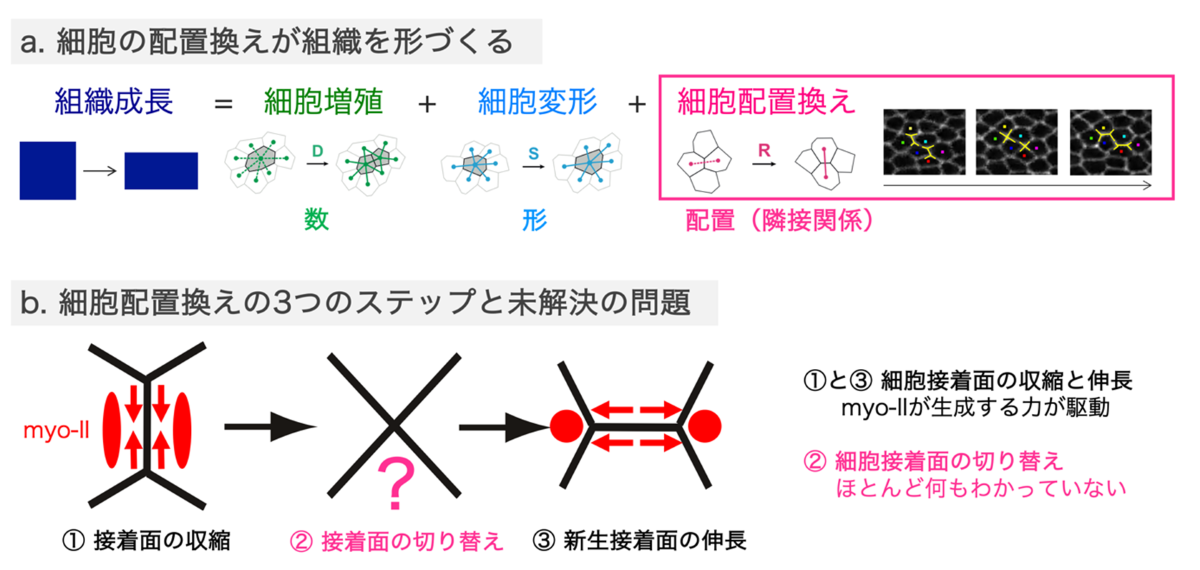
図1:組織の変形を駆動する細胞の配置換え
a.組織の変形は、細胞の数の変化(細胞分裂と細胞死)、細胞の形の変化、細胞の配置の変化に分解される。
b.細胞配置換えは、細胞接着面の収縮、切り替わり、伸長の三段階で進行する。
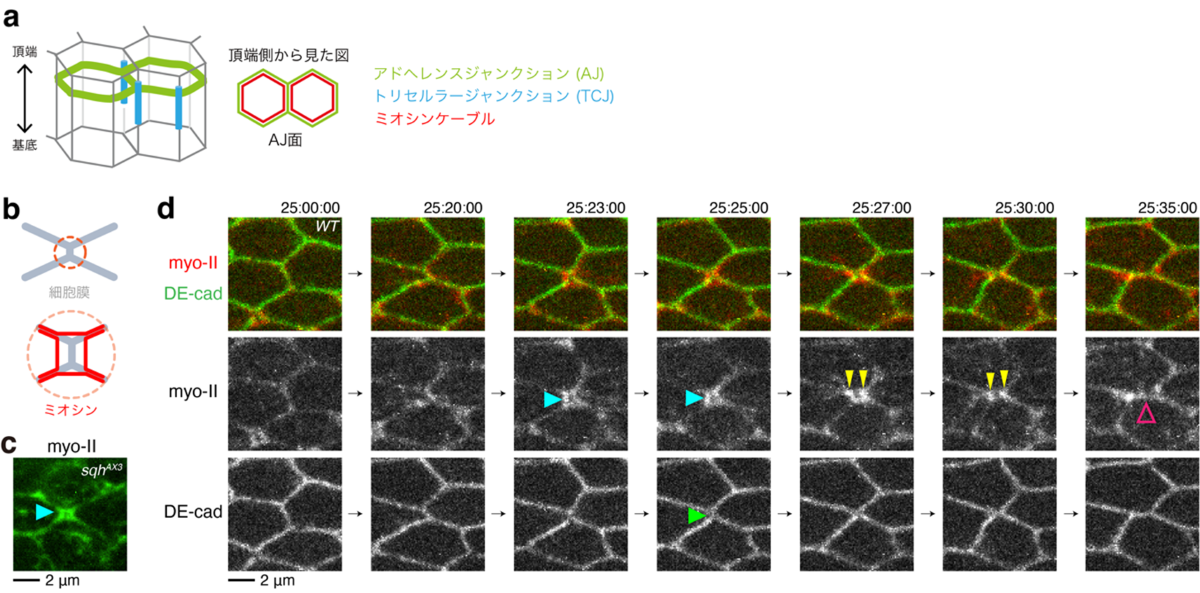
図2:rectangle-shaped myo-II cable (rsMC) の一過的な形成
a.上皮細胞の模式図。アドへレンスジャンクション(緑)にそってミオシンケーブル(赤)が走る。三つ以上の細胞が接着する点にトリセルラージャンクション(青)が形成される。
b.rectangle-shaped myo-II cable (rsMC) の模式図。上図のオレンジで囲った領域を拡大して下図に示す。赤:ミオシンケーブル、灰色:細胞膜。
c.ショウジョウバエ翅上皮細胞のrsMCの画像。緑:myo-II(ミオシン-II)。
d.rsMCの動態を可視化したタイムラプス画像。青矢頭はrsMCの形成、黄色矢頭はrsMCの分裂、マゼンタ矢頭はミオシンのアドヘレンスジャンクションへの再接着を指す。緑:DE-cadherin、赤:myo-II。
[研究内容]
本研究ではまず、rsMCの動態を高解像度タイムラプスイメージングにより追跡しました(図2d)。その結果、rsMCが細胞接着面の切り替わり時に一過的に形成されることがわかりました。rsMCの制御因子を探索したところ、アドへレンスジャンクションとアクチンの間のリンカー分子として働くことが知られているJub/Ajubaの発現を低下させると、rsMCが通常よりも早い段階で形成され、細胞配置換えの方向性が乱れることが明らかになりました。次に、野生型細胞におけるJub/Ajubaの局在を観察したところ、細胞接着面の切り替わり時に一過的に、Jub/Ajubaの局在が減弱していました。Jub/Ajubaの局在を制御する分子を探索する過程で、一部のトリセルラージャンクションからミオシンが剥離していることを見出したことから、トリセルラージャンクション構成因子の機能欠損実験を実施しました。その結果、M6と呼ばれる四回膜貫通型タンパク質がJub/Ajubaの局在を減弱させることがわかりました。さらに、Jub/AjubaがM6をアドへレンスジャンクションから排除していることが明らかになりました。以上から、①細胞配置換え過程で細胞接着面が収縮して、トリセルラージャンクション(細胞頂点)が近接すると、M6のJub/Ajubaへの阻害効果が強まり、Jub/Ajubaの局在が減弱して、rsMCが形成される、②細胞接着面が切り替わり、新しくできた細胞接着面が伸長すると、M6のJub/Ajubaへの阻害効果が弱まるので、ミオシンがアドへレンスジャンクションに再接着することが示唆されました(図4)。
ミオシンケーブルの物理特性がミオシンの細胞接着面からの剥離と再接着を制御する仕組みを明らかにするために、濡れの理論(注4)に基づいた力学モデルを構築しました(図3)。このモデルでは、ミオシンケーブルの張力と細胞接着面との接着による自由エネルギーから、ミオシンの剥離と接着が決定されます。モデルの解析から、接着状態から剥離状態へのエネルギーバリアが存在すること、ミオシンケーブルの張力がある大きさを超えると剥離状態に遷移することが示されました。興味深いことに、エネルギーバリアと張力の閾値は共に、細胞接着面の長さが短くなると小さくなる、すなわち、剥離状態がエネルギー的に好まれることがわかりました。さらに、細胞配置換え過程で観察されるJub/Ajubaの局在の減弱とミオシンの濃縮が、エネルギーバリアと張力の閾値を小さくする方向に働くことも明らかになりました。このように、細胞の力学とシグナリングが接着面の幾何特性と連関することで、細胞配置換えにおける細胞接着面の収縮と伸長に伴って、自発的に、細胞接着面の切り替えが起こると考えられます。
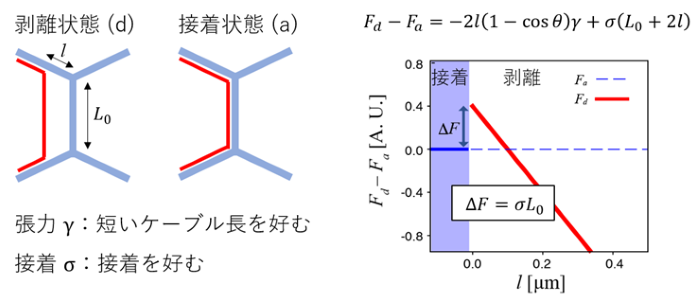
図3:ミオシンケーブルの張力と接着に由来する自由エネルギーに基づく力学モデル
ミオシンケーブルは張力を生成し、Jub/Ajubaなどのリンカータンパク質を介してアドへレンスジャンクションと接着する。前者と後者の競合により、剥離と接着のどちらの状態が実現されるかが決定される。剥離状態と接着状態のそれぞれについて自由エネルギーを算出し、その大きさを比べることで、剥離が起こる条件を求めた。エネルギーバリア (∆F) と剥離状態と接着状態のエネルギー差 (Fd-Fa) は、張力パラメータγや接着パラメータσに加えて、細胞接着面の長さや角度にも依存する。ミオシンケーブルの張力増加と接着減弱に加え、細胞接着面が短くなることがミオシンケーブルのアドへレンスジャンクションからの剥離を促す。
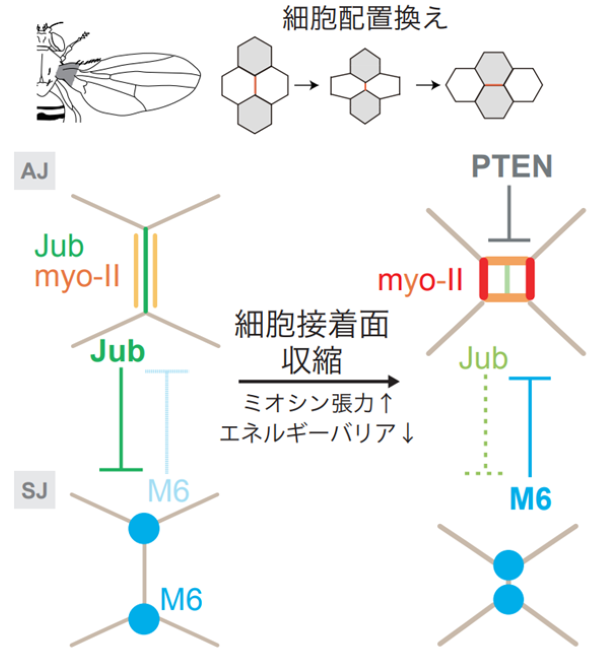
図4:本研究の概要
本研究ではショウジョウバエ翅上皮を実験系に用いて、細胞接着面の切り替わりのメカニズムを解析した。Jub/Ajubaはアドへレンスジャンクション (AJ) 面に、M6はアドへレンスジャンクション (AJ) 面よりも基底側のセプテートジャンクション (SJ) 面のトリセルラージャンクション(細胞の頂点)に局在する。細胞配置換えの初期段階において、細胞の頂点に局在するM6は、Jub/Ajubaに対して強い抑制効果を発揮できない。そのため、Jub/Ajubaはミオシンケーブルとアドヘレンスジャンクションの接着を維持することができる。細胞の配置換えが進むと、細胞接着面の収縮に伴って、細胞の頂点が近接し、M6のJub/Ajubaに対する抑制効果が高まり、Jub/Ajubaの局在が減弱し、rsMCが形成される。細胞接着面が切り替わって、新しくできた細胞接着面が伸長すると、M6のJub/Ajubaに対する抑制効果が再び弱まり、ミオシンケーブルが細胞接着面に再接着する。PTENは、翅の遠近軸方向(水平方向)のrsMCの辺におけるミオシンを抑制することで、遠近軸方向の細胞接着面へのミオシンの再接着を促進する。
[今後の展開]
細胞間接着の動態は生き物の体の維持から老化などのライフイベント、がんなどの病気と深く関わっています。本研究により明らかにされた細胞間接着制御メカニズムは、組織形態形成のみならず、さまざまな生命現象において働いている可能性があります。
本研究は以下の支援を受けて実施されました。
日本学術振興会科学研究助成事業(17K15125)、日本学術振興会研究助成事業(19K16139)、日本学術振興会研究助成事業(18KK0234)、公益財団法人住友財団助成金 (200303)、公益財団法人武田科学振興財団助成金、公益財団法人上原記念生命科学財団助成金 (202110172)、日本学術振興会国際共同研究事業 スイスとの国際共同研究プログラム(JPJSJPR20191501)、国立研究開発法人日本医療研究開発機構の革新的先端研究開発支援事業 (20gm5810025h9904)、科学技術振興機構戦略的創造研究推進事業(JPMJCR1923)
発表雑誌
雑誌:Current Biology
論文タイトル:Attachment and detachment of cortical myosin regulates cell junction exchange during cell rearrangement in the Drosophila wing epithelium
著者:Keisuke Ikawa*, Shuji Ishihara, Yoichiro Tamori, Kaoru Sugimura*
DOI番号:https://doi.org/10.1016/j.cub.2022.11.067
用語説明
(注1)ミオシン
分子モーターの一つ。アクチン細胞骨格とともにアクトミオシンを形成し、収縮力を生成する。アドヘレンスジャンクションを裏打ちするアクトミオシンは、細胞接着面を収縮させて、細胞の配置換えを駆動することが知られている。
(注2)アドヘレンスジャンクション
上皮細胞同士の接着を担う構造。膜貫通型タンパク質であるカドヘリンが主要な接着分子として働く。カドヘリンの細胞内ドメインは、Jub/Ajubaやα-cateninなどのリンカータンパク質を介して、アクチンと結合している。
(注3)トリセルラージャンクション
三つ以上の細胞が接着する細胞間接着構造。アドヘレンスジャンクションのような二細胞間の接着を担う構造とは構成分子が異なる。
(注4)濡れの理論
異なる媒質間の表面張力・界面張力のバランスによって、液体の濡れやすさ(親水性)や濡れにくさ(撥水性)、接着性を記述する理論。
[発表主体:東京大学 大学院理学系研究科]
―東京大学大学院総合文化研究科・教養学部 広報室―

