ニュース
![]() 2025.04.11
2025.04.11
【研究成果】光でゲノム変化を制御するゲノム合成技術「MagTAQing」を開発
2025年4月11日
東京大学
科学技術振興機構(JST)
発表のポイント
- 生物の性質を効率的に改良できるゲノムの再編成誘発技術TAQingシステムを改良し、青色光でゲノム変化を制御できるMagTAQingを開発しました。
- 青色光の下で活性化し、DNAを切断する制限酵素を世界で初めて開発しました。これを細胞内に導入したのち、細胞に青色光を照射すると、DNAが切断されるようになり、ゲノムの再編成を誘発することができました。
- 本技術により、細胞に与えるダメージを最小限にしつつ、ゲノム改変のパターンやレベルを自在に制御できるほか、狙った細胞や器官でゲノムの変化を誘発させることが可能になります。
概要
東京大学大学院総合文化研究科の太田邦史教授らの研究グループは、光で制御するトップダウン型ゲノム合成技術(注1)の開発に成功しました。
生命の設計図であるゲノム(注2)DNAを改変・合成し、医療や環境などの地球的課題を克服する新しい生物機能の開発が世界的に研究されています。このような技術をゲノム合成といいます。その中で、既存のゲノムDNAを再編成して生物機能を改良するリフォーム型の方式をトップダウン型ゲノム合成と呼びます。この方法は古くから交配や突然変異誘発によって人類が用いてきた生物改良法ですが、近年ではゲノムの再編成を誘発する手法が開発されています。
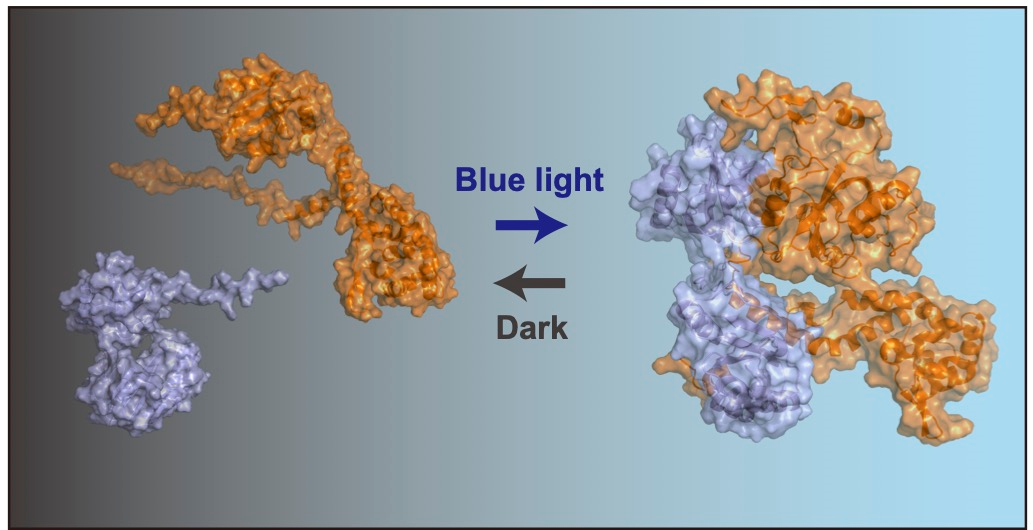
本研究では、東京大学で既に確立した技術であるTAQingシステム(注3)を改良し、青色光を照射したときのみゲノム再編成を誘発する改良技術MagTAQingを開発しました。重要な改良点としては、DNA切断酵素である制限酵素(注4)を最適な部位で分割し、その断片それぞれに青色光下で会合可能なマグネット(注5)というタンパク質を連結させ、青色光を照射したときのみ活性化する制限酵素を初めて創り出した点です。この技術を用いることで、MagTAQingを導入した細胞で、青色光を照射した時のみDNA切断を誘発でき、ゲノム再編成の発生を時間・空間的に精密制御できるようになりました。
発表内容
何を解決したか:
古来人類は、生物自身が持つDNA再編成のしくみ(交配による両親DNAの組換え)や突然変異処理(放射線照射や薬剤処理)を利用して、有用な性質を持つ品種を取得してきました。これらの方法では、目的の性質の生物を得るには非常に長い期間がかかります。一方、TAQingシステムでは、短期間で様々な形質を有する生物を得ることが可能です(Muramoto et al., Nature Commun. 2018)。当初開発されたTAQingシステムでは、温度で活性を制御可能な制限酵素を細胞内に導入し、一次的に加温して部分的にDNA切断活性を活性化させ、ゲノムDNAの再編成を誘発していました。ところが、温度変化に弱い生物や細胞への適用が困難であり、また特定の時期に狙った細胞だけDNA切断を誘発することは困難でした。今回、光で活性を制御できる制限酵素を新たに開発し、光で精密にDNA切断を制御することで、これらの課題を克服しました。
どう解決したか:
共同研究者の佐藤守俊教授(総合文化研究科広域科学専攻)の研究室で開発されていたマグネットタンパク質タグを様々なパターンで分割した制限酵素MboI(4塩基認識であるため、ゲノム中に非常に多くの切断部位を有する)の断片に連結し、酵母細胞内で発現させました(図1左)。この酵母細胞を青色光照射した場合としない場合で生育を比較し、青色光下でのみ細胞の生育が見られないMagMboI-8の組み合わせを選別しました。MagMboI-8を持つ細胞に青色光をあてると、マグネットタンパク質タグの会合により、制限酵素活性が発揮される正しい三次元構造が出来ることで、確かにゲノムDNAが細胞内で切断されていることを確認しました(図1右)。
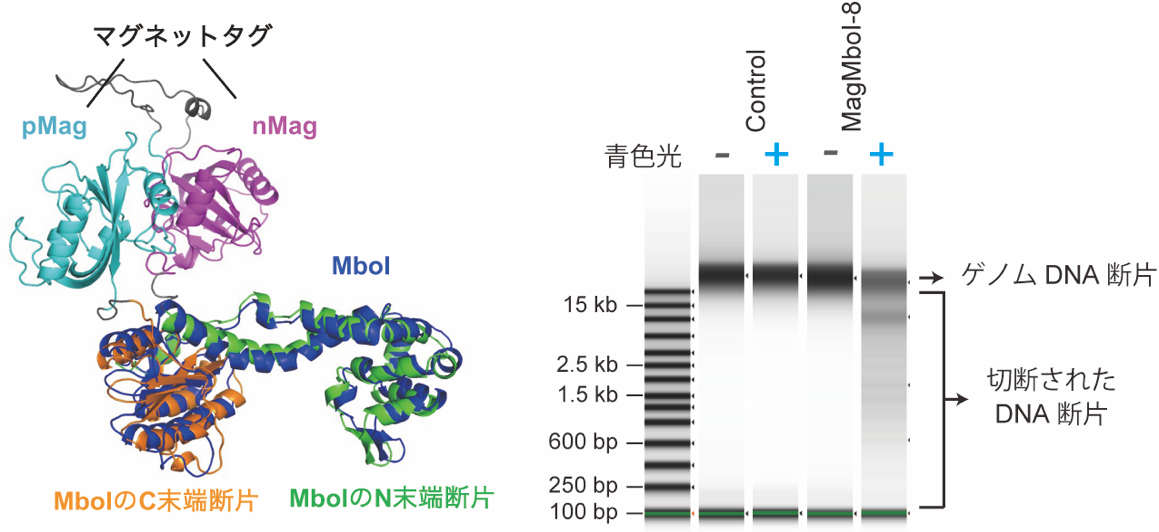
(右)青色光照射依存的なMagMboI-8のDNA切断活性。MagMboI-8 を発現した右端の試料だけ、DNAが分解して分子量の低い位置に尾を引いたように多数のDNA断片が見られる。
青色光を照射した細胞を分離し、全ゲノムDNA配列を解析した結果、多様なゲノム再編成が生じていることが確認できました。一部の染色体組換え部位では、MboIの認識配列の箇所で染色体が入れ替わっていること(転座)も確認できました(図2)。
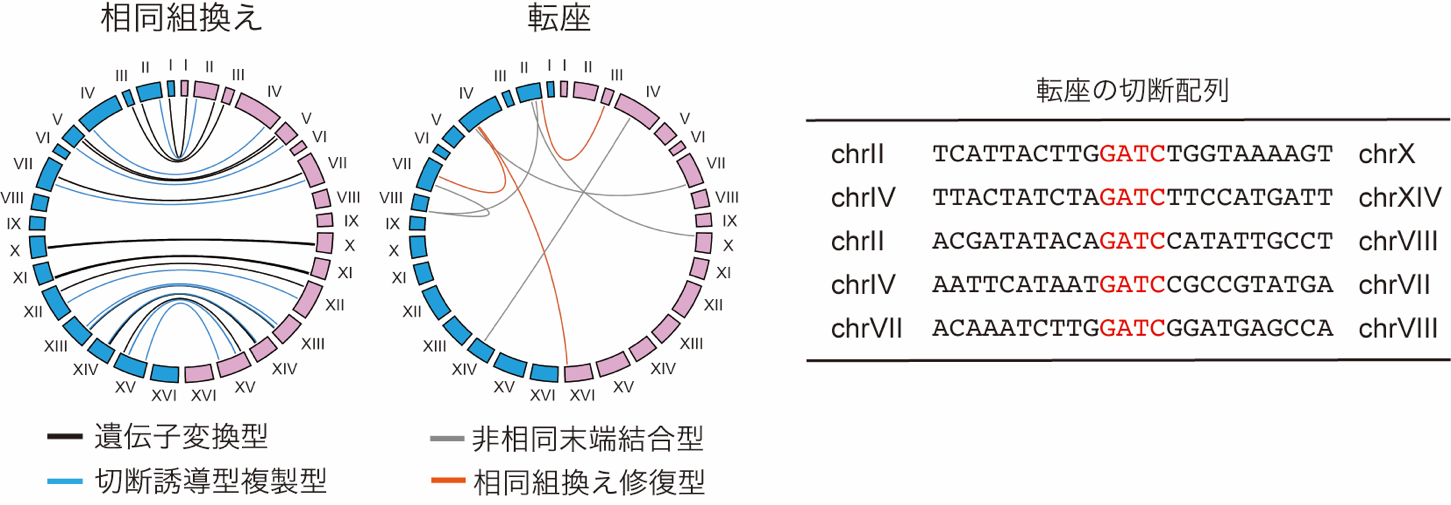
(右)非相同末端結合型転座の切断配列の例。MboIの認識配列(5'-GATC-3')を介して再結合されたことが確認された。
光で制御可能なゲノム再編成系に関する応用例として、温度変化に弱い細胞を用いた実験を行いました。減数分裂(注6)期の酵母細胞は温度上昇に弱いことが知られています。そこで、MagMboI-8を減数分裂期の酵母細胞に導入する実験を行いました。その際、減数分裂期組換えを担うSpo11(注7)というDNA切断酵素を欠損させた酵母細胞を用いました。なお、太田研究室ではこれまでに、Spo11を欠損する酵母細胞では減数分裂期組換えが全く起きないことを確認しています(Kawashima et al., Genes Cells, 2023)。
MagMboI-8を導入した減数分裂期のSpo11欠損酵母では、青色光照射で染色体組換えが多数誘発できることがわかりました(図3上段)。また、Spo11で生じる組換えでは見られない高頻度の染色体交換や、通常組換えが抑制されている組換え抑制領域(セントロメア近傍(注8)やリボゾーマルDNA領域(注9)など)でも高頻度で組換えが生じることがわかりました(図3下段)。温度上昇が必要な従来のTAQingシステムでは出来なかった、交配による遺伝子改良のハードルとなっている組換え抑制領域でのゲノム再編成が、今回の光制御型MagTAQingによって可能になり、品種改良の新たな可能性が拡大されたことになります。一方で、MagMboI-8を導入したSpo11欠損株では、生存可能な配偶子の頻度がかなり低いことがわかりました。この結果から、Spo11を介した自然界の染色体組換えが、子孫に継承する染色体の異常を抑制する機能を有することも明らかになりました。
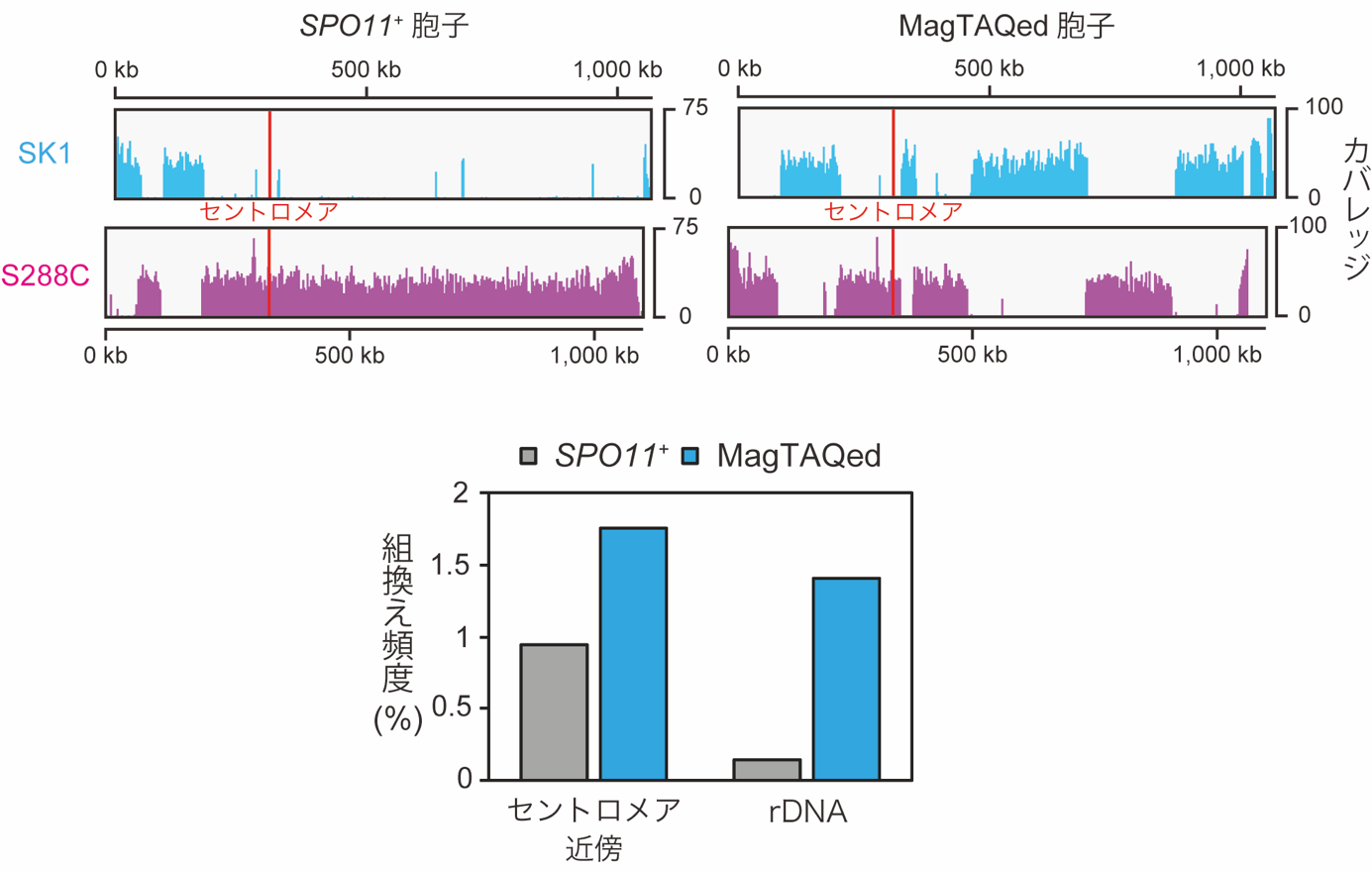
(下段)セントロメア近傍およびリボゾーマルDNA(rDNA)領域における組換え頻度。
発表者・研究者等情報
東京大学大学院総合文化研究科 広域科学専攻
米 秀之 特任研究員
川島 友莉 研究当時:博士課程
太田 邦史 教授/東京大学生物普遍性連携研究機構 教授/東京大学微生物科学イノベーション連携研究機構 教授
佐藤 守俊 教授
論文情報
雑誌名:「Nucleic Acids Research」(オンライン版)
題名:Light-controlled Spo11-less meiotic DNA breaks by MagTAQing lead to chromosomal aberrations
著者名:Hideyuki Yone†,*, Yuri Kawashima†, Hayato Hirai, Arisa H. Oda, Moritoshi Sato, Hiromitsu Kono, and Kunihiro Ohta* († 共同筆頭著者、*責任著者)
DOI:10.1093/nar/gkaf206
研究助成
本研究は、科学技術振興機構(JST) 戦略的創造研究推進事業CREST(課題番号:JPMJCR18S3)、 同 次世代研究者挑戦的研究プログラムSPRING(課題番号:JPMJSP2108)、革新的先端研究開発支援事業「感染症治療薬の新モダリティー天然物2.0」(課題番号:JP22gm1610007)、科学研究費補助金(課題番号:24K02068ST)、の支援により実施されました。
用語説明
(注1)トップダウン型ゲノム合成
既存のゲノムDNAを再編成し、生物の機能を改良する技術です。従来の交配や突然変異誘発に加え、近年では人工的にゲノム再編成を誘発する方法が発展しています。
(注2)ゲノム
ある特定の生物種を記述する最小単位のDNA情報です。細胞一つ一つにゲノムの情報をもつDNA(ゲノムDNA)が格納されています。
(注3)TAQingシステム
高度好熱菌(Thermus thermophilus)由来の制限酵素TaqIを使って、大規模なゲノム再編成を誘導する技術です。TAQingシステムでは、まずTaqIタンパク質を細胞内で発現させ、一時的に加温することで、DNA切断活性を活性化させます。活性化されたTaqIは、DNA切断を同時多発的に誘導し、DNA修復を介したゲノム再編成がランダムに誘発されます。TAQingシステムを酵母や植物に導入することで、エタノール発酵能が向上した酵母株やバイオマスが増大した植物株を取得することにも成功しています。
(注4)制限酵素
4塩基あるいは6塩基、またはそれ以上の長さのDNA塩基配列を特異的に認識してDNAを切断する酵素です。一部のバクテリアに存在し、バクテリアに感染するファージの増殖を制限する働きがあるため、制限酵素と呼ばれるようになりました。制限酵素は決まった配列でDNAを切断する性質があるため、組換えDNA技術の発展に不可欠な貢献を果たしました。
(注5)マグネット
アカパンカビ(Neurospora crassa)由来の光受容体タンパク質 Vividを改良して開発された青色光下で会合する一対の光スイッチタンパク質です。他の光受容体と比べ分子量が小さく、秒単位の速度でスイッチオン・スイッチオフできることや、光受容するための特別な補因子が不要であることが利点です。
(注6)減数分裂
性を有する生物は子孫への遺伝情報継承のため、配偶子(精子や卵)を作る際に減数分裂という特別な方式で細胞分裂を行います。減数分裂では1度のDNA合成の後、2回の連続した分裂を経て、ゲノムDNAが半分に減少します。第一減数分裂では、両親由来の相同染色体が対合した後に染色体の部分的な切断・再結合(遺伝的組換え)が高頻度で生じます。その後の第二分裂では、DNA複製で生じたコピーの染色体(姉妹染色分体)が分離し、それぞれ独立した配偶子へと分配されます。このプロセスによって、子孫の遺伝的な多様性が創出されます。
(注7)Spo11
酵母からヒトまで広く保存された減数分裂に特異的に出現するDNA切断酵素です。Spo11によるDNA二本鎖切断は減数分裂組換えの開始に必須であり、Spo11欠損株では組換えが全く起きず、多くの場合配偶子が機能しなくなります。Spo11がDNAを切断するには、出芽酵母の場合、9つの補助因子が必要であり、その他にヒストン修飾や染色体の高次構造などの多層的な制御が関与しています。
(注8)セントロメア領域
細胞核を有する生物(真核生物)の染色体は線状で、細胞分裂時に紡錘体の微小管が結合し、染色体を分離する際の土台となる特別な染色体領域があり、セントロメアと呼ばれます。多くの真核生物ではセントロメア領域は繰り返し配列などが存在し、特別なタンパク質が結合しています。減数分裂期の組換えがセントロメア領域では抑制されることが知られています。
(注9)リボゾーマルDNA領域
細胞内のタンパク質合成を司るリボソームを構成するRNAの遺伝子が存在する領域です。この領域では、リボソーム遺伝子が繰り返し多数並んでいます。この領域でも減数分裂期組換えが抑制されることがわかっています。

